Kuidas loomad tajuvad maailma?

Meile on ammu teada viis põhilist meelt: nägemine, kuulmine, haistmine maitsmine ja kompimine. Peale selle on eri liiki loomadel spetsiaalsed meeleelundid, mida avastatakse järjest uutel loomarühmadel. Eelkõige võib nimetada võimet tajuda elektrivälja ja Maa magnetvälja; üha rohkem liike kasutab eri eesmärkidel kajalokatsiooni.
Seega on loomadel piisavalt võimeid, tajumaks seda maailma, kus nad elavad.
Ent tuleb tõdeda, et kuigi meeleelundite ehitust, nii-öelda riistvara, on juba küllaltki põhjalikult kirjeldatud, jagub selles vallas veel pikaks ajaks avastamisrõõmu, kuna selgroogseid loomi on maailmas arvukalt, umbes 70 000 liiki.
Märksa halvemini on teada tajumise tarkvaraosa ehk kuidas meeleelundite edastatud informatsiooni analüüsitakse. Mõistagi on välja selgitatud need ajuosad, kuhu asjaomased närvid suubuvad, samuti see, et saadud infot võrreldakse eri ajuosades varem salvestatud teabega, aga edasine on teada ainult üldjoontes.
Püüame saada ülevaate selgroogsete loomade meelelunditest ja veidi ka viisidest, kuidas nad neid kasutavad (jätame välja kalad, kes väärivad eraldi käsitlust). Kõigepealt vaatleme, kuidas loomad koguvad visuaalset informatsiooni.
Alustame imetajatest.
Eeldan, et imetajate silma üldehitus on lugejatele tuttav, seetõttu pakuvad ehk rohkem huvi mitmesugused eluviisiga seotud kohastumused. Imetajate silmade ehitus on enamasti küllaltki tüüpiline ja tihti on haistmismeel ja kuulmine neile tähtsamad kui nägemine.
Kuna evolutsiooniliselt on imetajad esialgu olnud uru- ja videvikuloomad – paljud liigid on seda praegusajalgi –, pole neil olnud erilist vajadust värve eristada, kuid hea valgustundlikkus on suuresti abiks nii kiskjatele kui ka nende saakloomadele. Sellele aitab kaasa valgust peegeldav läikepiige (tapetum lucidum), mis paikneb võrkkesta taga ja peegeldab tagasi võrkkesta juba läbinud valguse (valgustundlikud kepikesed ja kolvikesed asuvad enamikul selgroogsetel võrkkesta põhjas).
Olenevalt imetaja eluviisist erineb kepikeste ja värvusnägemise eest vastutavate kolvikeste osakaal võrkkestas, seega ka värvinägemine ja vastu võetavad lainepikkused, näiteks näriliste silmad on tundlikud ka ultraviolettkiirguse lainepikkuste suhtes.
Erilisi silmade ehituse kohastumisi ei ole isegi veelise eluviisiga imetajatel, välja arvatud delfiinidel: neil on võime ahendada silmapupilli hästi ahtaks, et suurendada sügavusteravust samamoodi nagu toimib fotoaparaadi diafragma.
Urgudes elavate imetajate seas leidub seevastu nii kehva nägemisega kui ka täiesti pimedaid liike, nagu pimemutt Talpa caeca, kelle silmad on nahaga täiesti kaetud. Ja ega täielikus pimeduses nägemismeelel suurt tähtsust olegi, selle kompenseerivad hästi arenenud kompimis- ja haistmismeel.
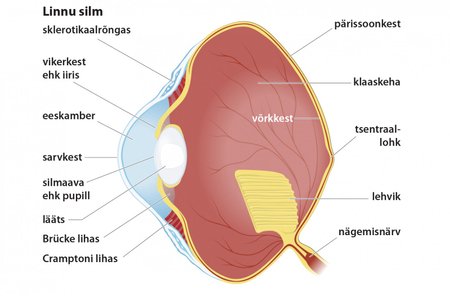
Lindudel on nägemismeel eluviisi tõttu esmatähtis meel ja väga hästi arenenud. Kehaga võrreldes on nende silmad väga suured ja paiknevad nii, et tagada võimalikult suur vaateväli. Silma niisutab ja kaitseb pilkkile.
Ehituse poolest erineb lindude silm mitmeti imetajate silmast. Põhierinevus on kahe fookustava lihase olemasolu: nõnda saab kiiremini fookustada. Kujutage ette, kui kiirelt peab linnu silm teravustuma, kui lendav pääsuke soovib napsata lendavat putukat!
Lindude silma klaaskehasse ulatub veresoonterikas kurruline lehvik, millel arvatakse olevat toitefunktsioon.
Kuna enamik linde tegutseb päeval, on neile oluline näha värvusi. Selleks on neil võrkkestas nelja ja mõnel liigil isegi viit tüüpi kolvikesi (imetajatel ainult kolm tüüpi). Kepikeste ja kolvikeste üldarv pindalaühiku kohta on ligi kaks korda suurem kui imetajatel ja mitut liiki lindude silmad näevad ka ultravioleti lainealas. Peale selle erinevad lindude kolvikesed ka ehituse poolest, näiteks võivad need sisaldada filtrina toimivaid õlitilgakesi.
Seega suudavad lindude silmad väga tõhusalt visuaalset infot koguda ja sedamööda on suurenenud ka keskaju nägemissagarad, mis on asjaomase teabe esmase töötlemise keskused.
Kahepaiksete ja roomajate silmad teravustatakse hoopis teisiti: muutes silmaläätse kaugust võrkkestast. Silmalääts ise on neil peaaegu kerakujuline. Puhkeasendis on kahepaiksete silmade on fookusse koondatud kaugel paiknevad objektid. Selleks et vaadata lähedale, liigub silmalääts vastava lihase abil võrkkestast eemale. Mitut liiki maod ja ka mõned kahepaiksed on võimelised tunnetama infrapunakiirgust, mida tajuvad retseptorid paiknevad madudel suuava ümbruses. Nõnda saavad nad tõhusalt jälitada ja tabada püsisoojaseid saakloomi.
Madude silmad toimivad samamoodi nagu kahepaiksetel, kuid teiste roomajate nägemiselund fookustab samal viisil kui lindude ja imetajate oma.
Siinkohal tasub mainida, et roomajatel ja ka kahepaiksetel võib olla ka kolmas, nn kiirusilm. Ehituselt sarnaneb see tavalise silmaga; arvatavasti on see seotud fotoperiodismiga.
Selgroogsete kuulmismeel.
Imetajatele on kuulmine üks tähtsamaid meeli, ent teistele selgroogsetele on see vähem oluline, aga maod on peaaegu kurdid. Kuulmismeel on aga oluline kajalokatsioonis; seda rakendavatel loomadel on kuulmiselund ka ehituse poolest iselaadne.
Usutavasti on kõrva ehitus üldjoontes teada, siin piirdume erisuguste kohastumuste ülevaatega.
Imetajate kõrv on võrreldes teiste selgroogsete rühmade omaga vast kõige efektiivsem helilainete vastuvõtmise organ. Erinevalt kõigist teistest rühmadest on imetajatel keskkõrvas kolm kuulmeluukest; trummikile ja sisekõrva ovaalakna pindalade erinevus suurendab kõrva tundlikkust helide suhtes. Kõrvalestad võimaldavad helisid veidi koondada ja tuvastada helide allika suuna.
Imetajate kõrva eripära seisneb veel selles, et umbes 18% imetajatel osaleb kõrv ka kajalokatsiooni käigus välja saadetud helisignaalide vastuvõtus. Peamiselt kasutavad kajalokatsiooni nahkhiired ja mitmed vaalalised, aga ka mõned teised imetajarühmad, nagu karihiired, pilukoonud ja tenrekid.
Õigupoolest avastati nahkhiirte kajalokatsioon juba 1793. aastal, kui Lazzaro Spallanzani tegi nende loomadega katseid.
Üldjuhul teatakse, et nahkhiired suudavad tekitada ja vastu võtta ultraheli, kuid paljud liigid kasutavad ka inimesele kuuldavaid sagedusi: nende sagedusala jääb enamjaolt vahemikku 12 kuni 100 kilohertsi. Seevastu vaalalised kasutavad suuremas osas inimesele kuuldavaid sagedusi. Erineb ka koht, kus heli tekib: vaalalistel ninaõõnes, enamikul nahkhiirtel neelupiirkonnas või tekitavad nad heli keelega. Ka signaali ribalaius ja modulatsioonitüüp on neil erisugused: nahkhiirtel ei ulatu kajalokatsiooni hõlmav ala üle 60 meetri (mõnel liigil ainult 2,5 m), vaalalistel küündib see kuni 1,5 kilomeetrini, mõnedel andmetel kuni 16 kilomeetrini.
Vaalaliste väljasaadetavad helid on väga valjud, näiteks ulatub kašelotil kuni 240 detsibelli – see on kõige tugevam looduslik heli. Selle põhjal on püstitatud hüpotees, et need helid mõjuvad paralüseerivalt nende saakobjektidele, tehes nad liikumisvõimetuks. Tõendusi selle kohta siiski veel ei ole.
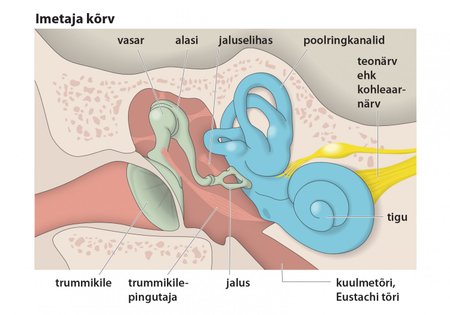
Kajalokatsiooni rakendavad imetajad peavad suutma vältida kurdistumist enda tekitatud helide mõjul. Selle tarbeks on neil mitu moodust. Näiteks nahkhiirtel on väga hästi arenenud kaks keskkõrvalihast: trummikilepingutaja ning jaluselihas, mis pingestuvad umbes kolm millisekundit varem kui heli tekitavad lihased. Nõnda väheneb tunduvalt kuulmeluude vibratsioon ja sedakaudu ka kuulmisteravus. Peale selle on nahkhiirtel kuulmepõiendid, kus paiknevad kesk- ja sisekõrv, mis on ülejäänud koljust eraldatud rasv- ja sidekoega täidetud urgete abil. Ka vaalalistel on kuulmepõiendid ülejäänud koljuluudest isoleeritud.
Delfiinide kajalokatsiooni võivad kuulda ka nende saakloomad. Ometi olid kaetud silmadega delfiinid võimelised tabama kogu saagi (100%) ilma nägemismeele abita ja ka kajalokatsiooni kasutamata. Nende tulemuste põhjal on järeldatud, et delfiinid on võimelised looma lähiümbrusest mürapildi. See on uus võimalus „näha häälega“.
Kindlasti on huvitav teada, et tuntud Eesti imetajatest võivad näiteks rändrott ja mõned uruhiireliigid tekitada ultraheli, kuid liigisisese suhtluse, mitte kajalokatsiooni tarbeks.

Lindudele on kuulmine küll vähem oluline kui nägemine, kuid siiski küllaltki hästi arenenud. Erinevalt imetajatest paikneb lindudel keskkõrvas ainult üks kuulmeluu; kõrvalesti neil ei ole, aga sama otstarbega moodustisi võib siiski leiduda, eriti kakulistel. Vasak ja parem kuulmekäik on lindudel eri pikkusega: närvisüsteemi abiga annab see neile veidi parema võime eristada helide suunda.
Laias laastus on lindude kuulmisulatus enam vähem sama mis inimesel ehk 20 – 20 000 hertsi. Seega ei kuule linnud ultraheli, kuigi mõned pimedas koobastes elutseval liigid võivad kasutada kajalokatsiooni.
Lindudele oluline sagedusala jääb vahemikku üks 1000–4000 hertsi, aga tuvid võivad kuulda ka poolehertsist madalsagedusheli. Arvatavasti suudavad nad seetõttu kuulda läheneva tormi heli ja teisigi looduses levivaid ülimadalsageduslikke signaale ning kasutada seda võimet ka navigeerides.
Seega võib öelda, et kuulmine on lindudel hea, mõnel liigil isegi väga hea.
Kahepaiksete kuulmismeel on samuti väga hea.
Mõnedel salamandritel leidub peale tavalise, õhu kaudu levivate võngete juhttee veel teine juhttee, et võtta vastu maapinna madalsageduslikke võnkeid. Enamasti võtavad kahepaiksete papillid – võimalikud imetajate kuulmisrakkude ehk Corti elundi eelkäijad – vastu õhu kaudu levivaid kõrgemaid sagedusi. Ent mõnel kahepaiksel võib sisekõrvas asuda veel teinegi papill (papilla amphibiorum), mis võtab vastu maapinna kaudu levivaid madalsagedusvõnkeid. Need võnked võivad kahepaiksetele anda teada vaenlase lähenemisest.
Roomajate kõrvas on samuti ainult üks kuulmeluuke ning enamikul on ka trummikile ja väline kuulmekäik. Seega, valdavalt on neil küllaltki hea kuulmismeel. Erand on maod, keda kaua aega peeti täiesti kurtideks, kuna neil puudub nii trummikile kui ka väliskõrv. Nimelt ei toetu nende sisekõrvas olev kuulmetulbakene mitte vastu trummikilet, vaid vastu üsna kogukat ruutluud, mis seob alalõualuu koljupõhimikuga. Katsete põhjal maod siiski reageerivad maapinna võngetele ja mingil määral ka õhuvõngetele, kuigi vastuvõetavate helide ulatus on üsna piiratud.
Haistmismeel ei ole käsitletavatele loomarühmadele ühtviisi tähtis.
Teadaolevalt on enamjaole imetajatele väga oluline haista, lindudel ei etenda see meel üldiselt kuigi suurt osa, seevastu paljudele raipetoidulistele lindudele ja kiivile on haistmine üsna tähtis. Kiivi on ainus linnuliik maailmas, kelle ninasõõrmed paiknevad noka otsas.

Siinkohal peame rääkima ka niinimetatud vomeronasaalorganist ehk Jacobsoni elundist, mida saab käsitada feromoonide tuvastamisele spetsialiseerunud lisahaisteelundina. See on olemas enamikul imetajatel, kahepaiksetel ja roomajatel. Selle abil teevad isasloomad näiteks kindlaks emaslooma valmisoleku sigida. Kui jäär läheneb emasele lambale ja tõstab ülamoka üles, siis üritab ta kindlaks määrata, kas konkreetne emasloom indleb: ülamoka all asetseb Jacobsoni organil ava, kustkaudu siseneb õhk. Ka roomajate ja kahepaiksete sigimiskäitumises on Jacobsoni organil oluline osa.
Käsitleda maitsmismeelt haistmisest eraldi on loomariigi puhul vahel veidi kunstlik, sest veeluviisiga selgroogsetel võivad kemoretseptorid paikneda üle keha. Seega võivad nad n-ö maitsta kogu kehaga. Nagu üldiselt teada, eristavad meie suus olevad maitsepungad viit põhimaitset: magus, kibe, soolane, hapu ja umami (selle tekitavad glutamaadid ja nukleotiidid).
Loomade maitsmismeelt on kujundatud evolutsiooni vältel. Näiteks sõralistel, kelle maitsmismeel peab olema väga hea, eristamaks mürgiseid või söömiskõlbmatuid taimi, on suus kuni 25 000 maitsepunga, kuid kassil ainult 500. Inimene kui segatoiduline oma 10 000 pungaga jääb kuhugi keskele. Lindudel kipub neid olema õige vähe, kanal ainult 30. Paljudele loomadele on omased keriotstarbega maitsepungad, juhul kui nad on spetsialiseerunud selle maitsega toiduobjektile.
Kahepaiksete maitsmismeel on küllaltki spetsialiseerunud ja liigispetsiifiline. Mõru maitsega toitu siiski üldjuhuk ei tarbita, soolasega võib olla nii või teisiti. Sama kehtib ka näiteks sisalike kohta, kuigi nende maitsemeele kohta on vähe teada.
Kompimisaistinguid võtab õigupoolest vastu kogu keha. Vabad ja mitut tüüpi kapseldunud närvilõpmed paiknevad üle kogu keha ning ka karva- ja sulenääpsudes. Mõnes piirkonnas leidub neid rohkem, mõnes vähem. Lindudel on mõnda tüüpi suled ja imetajatel erilaadsed karvad spetsialiseerunud ainult kompeaistinguid vastu võtma, näiteks on öösorril noka ümber sulgedest koonus, mis annab linnule teada, et on paras aeg saak kinni napsata.
Lühidalt ka Maa magnetvälja tajumisest.
Katsetega on tõestatud, et kõikides käsitletud süstemaatilistes rühmades leidub liike, kes suudavad tajuda maa magnetvälja ja rakendada seda peamiselt orienteerumise otstarbel. Kahepaiksetest on sellise võime olemasolu kindlaks tehtud meilgi tavalisel kärnkonnal, roomajatest mitut liiki kilpkonnadel, lindudest tuvidel ja üllataval kombel koguni kanadel. Teada on isegi asjaomase sensori asukoht organismis, näiteks heinaturpialil paikneb retseptor (tsütokroom) silmas, kaart (magnetiit) aga kolmiknärvi silmaharus.
Mitu tavalist ja laialt levinud imetajaliiki on samuti võimelised tajuma magnetvälja, näiteks nahkhiired, rebane ja hirvlased. Paljudel juhtudel ei ole siiski suudetud magnetvälja suhtes tundlikke alasid või retseptoreid organismis usaldusväärselt tuvastada, kuna nende mõõtmed on väikesed.
Usutavasti võivad mõned tsütokroomid ja rauda sisaldavad struktuurid olla seda laadi retseptorite talitluse alus. Siiski on selles vallas veel palju uurida.
Võime tajuda elektrivälja on peamiselt kalade ja kahepaiksete pärusmaa, kuna vesi on hoopis parem elektrijuht kui õhk. Kuid seda suudavad siiski ka mõned teised veelised ja poolveelised selgroogsed. Põhiliselt on elektroretseptorid seda tüüpi nagu haidelt tuntud Lorenzini ampullid: elektrit juhtivad torukesed, mis viivad impulsid keha pinnalt vabade närvilõpmeteni. Imetajatest on kõnealune võime ainupilulistel (nokkloomal) ja teadaolevalt ka delfiiniliigil Sotalia guianensis. Nokkloomal asuvad ampullilaadsed struktuurid noka piirkonnas, toimivad antennidena ja võimaldavad tabada saaki piiratud nähtavusega veekeskkonnas.
Kõik eeltoodu annab tunnistust sellest, et teiste loomade suutlikkus tajuda maailma ei jää alla inimese omale: mõneti on nende meeled hämmastavalt arenenud; see on võimaldanud neil kohaneda ja jääda ellu üha kiiremini muutuvas maailmas.
Ent uurimistöö selles valdkonnas jätkub, kindlasti ilmneb palju uut teadaolevate meelte kohta ja võib-olla leitakse veel uusigi.
* Artikkel ilmus originaalis ajakirja "Eesti Loodus" oktoobrinumbris.