Haruldased kalifornia kondorid on hakanud järglasi saama ilma vastassoo abita
(9)San Diego loomaaed andis oktoobri lõpus teada, et sealsete kalifornia kondorite (Gymnogyps californianus) geeniuuringu käigus on selgunud erakordselt huvitav fakt. Kaks isaslindu, üks koorunud 2001. aastal ja teine 2009. aastal, on pärit seemnerakuga viljastamata munast. Praeguseks on mõlemad partenogeneetilised kalifornia kondorid surnud. Üks neist suri kahe aasta vanusena, teine elas seitsmeaastaseks.
Artikkel ilmus originaalis ajakirja Eesti Loodus detsembrinumbris.

Enamikul loomadel on suguliseks paljunemiseks vajalik, et sperm viljastaks munaraku. Isasloomad toodavad sperme ja emasloomad munarakke ning hermafrodiidid on võimelised tootma nii munarakke kui ka sperme. Selgrootutel (nt skorpionid, ümarussid, lestad, kiletiivalised jt) on küllaltki tavaline ka partenogenees ehk neitsissigimine, mille puhul embrüo areneb viljastamata munarakust. Hea näide on hästi tuntud kodumesilane (Apis mellifera), kelle viljastamata munadest arenevad haploidsed (ühekordse kromosoomiarvuga) suguvõimelised isased ehk leskmesilased. Selgroogsetel tuleb partenogeeneesi ette märgatavalt harvemini, üksnes mõningatel kahepaiksetel, kaladel, roomajatel ja lindudel.
Mitmel juhul on nende loomade partenogenees avastatud tehisoludes, mistõttu on see tekitanud omajagu furoori. Näiteks USA Chattanooga loomaaia (Tennessee osariik) emane komodo varaan Charlie oli paaris isase Kadaliga, kuid ilmselt vastupidi tolle ootustele munes
2019. aastal munad, millest koorus kolm partenogeneetilist isast Onyx, Jasper ja Flint. Louisville’i (Kentucky osariik) loomaaia emane võrkpüüton Thelma elas ühes puuris teise emasega, ja andis kõigile üllatuslikult partenogeneesi teel elu kuuele emasele järglasele.
Kõrgematel selgroogsetel on partenogeneesi kaudu arenema hakkavad embrüod elujõuetud, ehkki lindude puhul on harvadel juhtudel siiski tehtud kindlaks, et nad on võimelised kooruma ja arenema täiskasvanuks.
Imetajatel ei saa ainult emase genoomsest materjalist areneda eluvõimelist embrüot, kuna mängu tuleb epigeneetika. Nimelt on isas- ja emasgenoom erinevalt vermitud ehk imprinditud: geeni avaldumine oleneb sellest, kummalt vanemalt on see päritud. Osa arengus olulisi geene on ekspresseeritud isasgenoomilt ja osa emasgenoomilt. Kui näiteks eemaldada hiire viljastatud munarakust maternaalne (emapoolset päritolu) genoom ja asendada see teise paternaalse (isapoolset päritolu) genoomiga, saadakse alaarenenud embrüo, kes on määratud hukule (sama juhtub ka kahe maternaalse genoomi puhul).
Samamoodi juhtub siis, kui naise munaraku arengu käigus on maternaalne genoom meioosis kaduma läinud ja nii-öelda tühja munaraku viljastab kaks spermi. Selle tagajärg on arenguanomaalia, mida kutsutakse põismooliks. See on trofoblasti (põisloote välimise rakukihi / rakud, mis aitavad embrüol emakasse pesastuda, pannes aluse platsenta lootepoolsele osale) vohamine, kuna paternaalselt genoomilt ekspresseeruvad geenid, mis on vajalikud, moodustamaks platsentat.
Teatud juhtudel on molekulaarbioloogiliste võtetega võimalik ka imetajatel nii-öelda partenogenees esile kutsuda. Näiteks Jaapani teadlased lõid 2004. aastal hiire, kes oli saadud ilma isaslooma panuseta. Nimelt võeti mutantselt hiirelt pärit munarakk, mis oli muudetud imprintingu mustri mõttes spermilaadseks, kombineeriti see normaalse hiire munarakuga ja saadi lõpuks pärast mitut ebaõnnestumist elujõuline ja viljastusvõimeline emane hiir.
Ainsad selgroogsed, kellel teadaolevalt toimub genoomne imprinting, on pärisimetajad ja mõningal määral ka kukkurloomad. Munevatel ainupilulistel imetajatel ei ole imprinditud geene leitud. Lindude kohta arvatakse, et neil genoomset vermimist ette ei tule.

Roomajad: USA-s ja Mehhikos elutsev sisalik Aspidoscelis uniparens; komodo varaan (Varanus komodoensis); kuningboa (Boa constrictor); võrkpüüton (Malayopython reticulatus); vaskpea-kilplõugmadu (Agkistrodon contortrix).
Kõhrkalad: vasarhai Sphyrna tiburo; väikehammas-saagrai (Pristis pectinata); sebra-vööthai (Stegostoma fasciatum); mustserv-hallhai (Carcharchimus limbatus).
Varem dokumenteeritu järgi on embrüod moodustunud partenogeneetiliselt kodukanal (Gallus gallus domesticus), kodukalkunil (Meleagris gallopavo f. domestica), kodutuvil (Columba livia var. domestica), sebravindil (Taeniopygia guttata) ja ida-sinivutil (Coturnix chiensis). Üksnes vähestel juhtudel, kana ja kalkuni puhul, on tehtud kindlaks, et embrüod suudavad areneda kuni koorumiseni, ja kalkuni puhul on õnnestunud suunatud aretustööga saada isegi viljastusvõimelisi täiskasvanud isaseid. Miks just isaseid? Põhjus peitub sugukromosoomides.
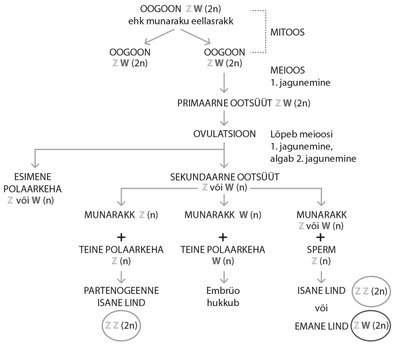
Lindude sugukromosoomid on Z- ja W-kromosoom. Emastel on Z- ja W-kromosoom, isastel kaks Z-kromosoomi. Seega, lindudel on emased heterogameetsed ehk toodavad nii Z- kui ka W-sugukromosoomiga munarakke. (Meeldetuletuseks: imetajatel, näiteks inimestel, on isased heterogameetsed, s.o toodavad X- ja Y-sugukromosoomiga sperme.) Kui emased ZW-genotüübiga linnud munevad viljastamata muna (kas Z-kromosoomi sisaldava muna või W-kromosoomiga muna), millest areneb partenogeneesi teel järglane, saab see olla ainult ZZ, seega isane, kuna WW-genotüübiga versioon pole elujõuline.
Normaalselt viljastudes siseneb isaslinnu sperm munarakku, munaraku meioos toimub lõpuni ning haploidne spermi (paternaalne) genoom ja haploidne munaraku (maternaalne) genoom ühinevad ning panevad aluse uuele diploidsele organismile. Partenogeneesis võivad munarakk ja meioosi käigus moodustunud teine polaarkeha taasühineda, tekitades diploidse kromosoomistiku. Kuna emane lind on ZW-genotüübiga, võivad munarakud ja polaarkehad sisaldada vastavalt kas Z- või W- sugukromosoomi. Kui nüüd ühinevad Z-kromosoomi kandev munarakk ja polaarkeha, on tulemus ZZ-genotüübiga isane embrüo, kelle genoomne materjal pärineb emalt. Kui ühinevad kaks W-kromosoomi kandvat rakku, siis embrüo hukkub.
Kalifornia kondori näide partenogeneesi võimalikkuse kohta lindudel on intrigeeriv, kuna loob võimaluse, et seda võib ka loodusoludes ette tulla märksa laiemalt, kui arvatakse. Samas näitas ka nende kahe partenogeneetiliselt arenenud isase kondori elusaatus, et selline arengumudel kaldub normist kõrvale ja põhjustab mitmesuguseid arenguhäireid (anatoomilised iseärasused, lühem eluiga, haigused, kaheldav sigimisvõime jne). Suure tõenäosusega hukkuvad looduslikult tekkinud partenogeneetilised linnud juba embrüogeneesi käigus ega kooru, aga välistada seda võimalust ei saa.
Mõlemad San Diego loomaaia emased kondorid, kelle munadest koorusid partenogeneetilised järglased, olid puuris koos isastega ja varem saanud elujõulisi tibusid suguliselt paljunedes. Seega võib sellist partenogeneesi pidada fakultatiivseks ehk suguliselt paljuneva organismi võimeks saada järglasi ka aseksuaalse paljunemise teel.